14 Apr 2021
The aim of vaccination is to induce immunity to a specific pathogen without causing disease.

When a vaccinated animal is exposed to natural challenge, antigen recognition triggers an immune response, which leads to the expansion of pathogen-specific memory B cell and T cell populations.
B cells go on to produce antibodies, which can neutralise the pathogen or aid pathogen clearance though phagocytosis. T cells either coordinate the immune response (cluster of differentiation [CD]4+ helper T cells) or recognise and destroy infected cells (CD8+ cytotoxic T cells). This enables the animal to clear the infection before the pathogen can cause disease.
For vaccines to provide optimal protection, they need to:
As bovine respiratory disease (BRD) viruses initiate infection within the respiratory tract, inducing immunity at the mucosal surfaces can optimise protection against infection – mucosal antibodies block virus entry into cells, while cytotoxic T cells kill infected cells.
The mucosal immune system is distinct from the systemic immune system. It provides a first-line defence against pathogen invasion at mucosal surfaces, mediated by aggregates of immune cells located near to the mucosa. This is known as mucosa-associated lymphoid tissues (MALT).
The most effective way of inducing mucosal immune responses is by administration of vaccines on to the mucosal surface.
Antigen delivered to the mucosal surface is primarily taken up by M cells, or goblet cells, and delivered to dendritic cells (DC) underlying the mucosal epithelium. These DCs then present this antigen to CD4+ and CD8+ T cells in MALT or mucosal lymph nodes.
Antigen also travels directly to the lymphoid tissue, where it is recognised by antigen-specific B cells.
A subset of CD4+ helper T cells migrate to B cell follicles within the lymphoid tissue to help B cells produce high-affinity antibodies. B lymphocytes and T lymphocytes then exit the mucosal lymphoid tissues and recirculate via the blood before re-entering mucosal tissues 7 to 10 days later as a result of specific homing receptors on their surfaces.
Once there, T cells and B cells exert their effector responses; B cells differentiate into plasma cells and produce IgA, which is secreted on to the mucosal surface and can block pathogen entry, and CD8+ cytotoxic T cells kill infected cells. This process is shown in Figure 1.
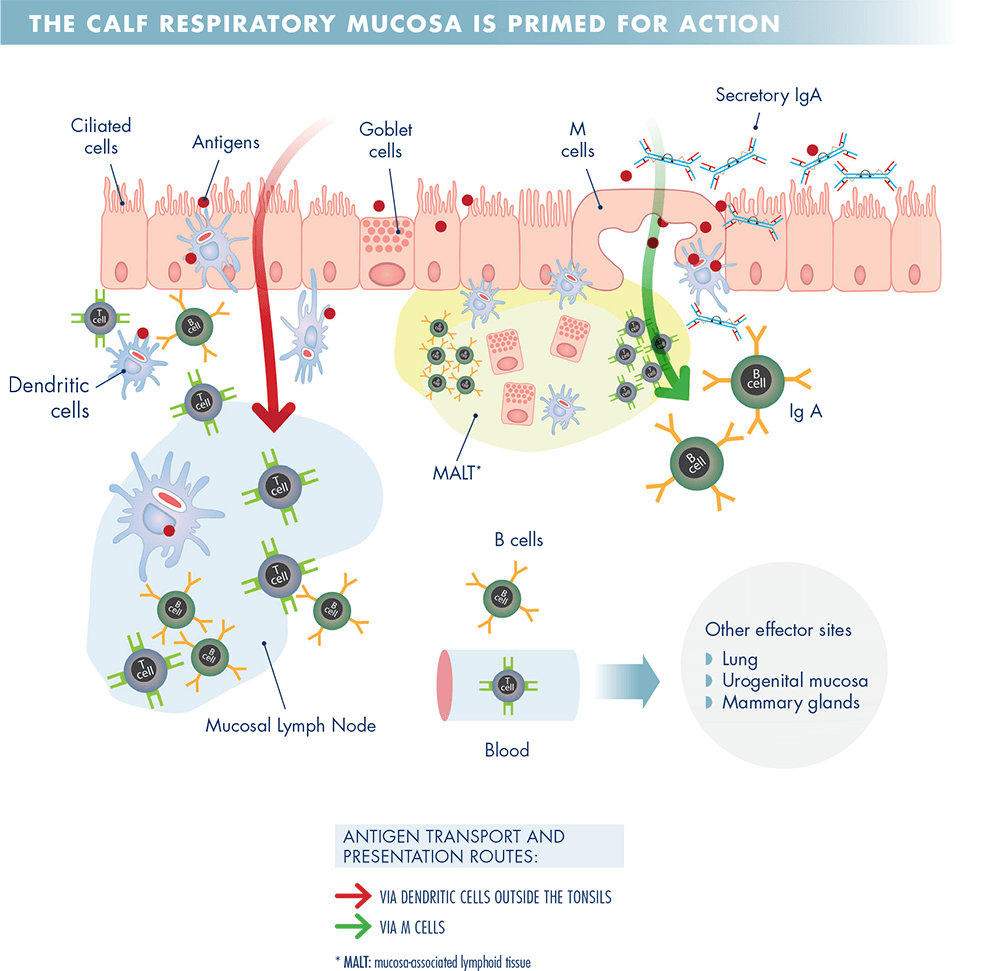
As severe BRD viral infections tend to occur in young calves, it is desirable that vaccines work in the face of maternally derived antibodies (MDA) and provide rapid immunity.
Binding of maternal antibodies to vaccine antigens can interfere with the response to systemic (injectable) vaccines. This inhibits B cell activation and attenuates the initial antibody response to the vaccination, although a B cell memory response may be induced, and T cell priming will still occur2.
Studies have shown that live attenuated viral vaccines delivered mucosally (intranasally) are more effective at inducing protection in the face of MDA than inactivated or live vaccines delivered by injection3. A recent study demonstrated that MDA-positive calves vaccinated with Bovalto® Respi Intranasal were protected against infection by challenge with either parainfluenza-3 or bovine respiratory cyncytial virus at 12 weeks after initial vaccination4.
The efficacy of mucosal vaccines in the face of MDA is due to the presence of lower levels of virus-specific maternal antibodies at the mucosal surface compared to SC or IM injection sites. Induction of non-specific cytokine-mediated innate immunity, and effective priming of cellular immune responses within the respiratory tract enhance their effect.
Mucosal immunity develops rapidly, but typically provides a shorter duration of immunity than that conferred by systemic administration of injectable vaccines. Follow-up vaccination can extend the duration of immunity against BRD pathogens and provide appropriate protection ahead of high-risk periods5.
BRD virus vaccines need to generate effective immune responses within the respiratory tract. Live attenuated vaccines delivered on to the respiratory tract provide an efficient means of achieving this. These vaccines induce both innate and adaptive immune responses that contribute to the protection against BRD viruses, and, importantly, can work in the face of MDA, allowing their use in young calves.
Bovalto® Respi Intranasal, nasal spray, lyophilisate and solvent for suspension contains bovine parainfluenza-3 virus, modified live virus, strain Bio 23/A 105.0 – 107.5 TCID50 and bovine respiratory syncytial virus, modified live virus, strain Bio 24/A 104.0 – 106.0 TCID50. Bovalto® Respi 3 Suspension for Injection and Bovalto® Respi 4 Suspension for Injection contain inactivated bovine respiratory syncytial virus, strain BIO-24, inactivated bovine parainfluenza-3 virus, strain BIO-23 and inactivated Mannheimia haemolytica, serotype A1 strain DSM 5283. Bovalto® Respi 4 also contains inactivated bovine viral diarrhoea virus, strain BIO-25. UK: POM-V. Advice should be sought from the prescriber.
Further information available in the SPCs or from Boehringer Ingelheim Animal Health UK Ltd, RG12 8YS, Tel: 01344 746957. Email: [email protected]
Bovalto® is a registered trademark of the Boehringer Ingelheim Group. ©2020 Boehringer Ingelheim Animal Health UK Ltd. All rights reserved. Date of preparation: Oct 2020. BOV-007-2021. Use medicines responsibly.